
Qu'est-ce qu'un dinosaure ? Cette question, qui semble pourtant tellement simple, est en fait beaucoup plus complexe qu'il n'y paraît... Comment, en effet, donner à cette question une réponse complète ? Comment être sûr de ne pas avoir omis un point essentiel mais pour l'instant encore inconnu ?
Les dinosaures forment un clade extrêmement diversifié de vertébrés diapsides (possédant deux ouvertures temporales). Ils présentent une grande diversité de tailles, de formes... et certainement de couleurs ! Ce sont des archosaures (qui comprennent également les crocodiliens et les oiseaux) ovipares, ayant en commun une posture érigée, avec les membres directement sous le corps, et un certain nombre de synapomorphies (caractère dérivé partagé par plusieurs taxons).
Les plus anciens fossiles connus de dinosaures remontent au début du Trias supérieur, il y a près de 230 millions d'années, à l'aube du Mésozoïque. Cependant, leur origine est sans doute bien antérieure. Présents sur l'ensemble des continents pendant plus de 160 millions d'années, ils disparaissent presque entièrement à la fin du Crétacé, il y a 65 millions d'années, suite à la combinaison de divers événements, dont la chute d'une météorite. Ne subsiste plus aujourd'hui qu'une infime partie de l'ancienne diversité de ces animaux : les oiseaux. Les dinosaures forment l'un des groupes de vertébrés les plus florissants, et les tétrapodes les plus diversifiés taxonomiquement parlant, le nombre de leurs espèces étant supérieur à celui des mammifères. A ce jour, environ 700 espèces de dinosaures non-aviens (exclusion faite des oiseaux, donc) ont été découvertes. Un peu plus de la moitié d'entre elles ne sont connues que par un seul spécimen, et moins de 20% sont connues par plus de cinq spécimens. Cependant, un étude récente suggère que 71% des genres de dinosaures restent encore à découvrir, ce qui porterait à environ 1850 le nombre d'espèces de dinosaures ayant existé.
Les dinosaures forment un clade extrêmement diversifié de vertébrés diapsides (possédant deux ouvertures temporales). Ils présentent une grande diversité de tailles, de formes... et certainement de couleurs ! Ce sont des archosaures (qui comprennent également les crocodiliens et les oiseaux) ovipares, ayant en commun une posture érigée, avec les membres directement sous le corps, et un certain nombre de synapomorphies (caractère dérivé partagé par plusieurs taxons).
Les plus anciens fossiles connus de dinosaures remontent au début du Trias supérieur, il y a près de 230 millions d'années, à l'aube du Mésozoïque. Cependant, leur origine est sans doute bien antérieure. Présents sur l'ensemble des continents pendant plus de 160 millions d'années, ils disparaissent presque entièrement à la fin du Crétacé, il y a 65 millions d'années, suite à la combinaison de divers événements, dont la chute d'une météorite. Ne subsiste plus aujourd'hui qu'une infime partie de l'ancienne diversité de ces animaux : les oiseaux. Les dinosaures forment l'un des groupes de vertébrés les plus florissants, et les tétrapodes les plus diversifiés taxonomiquement parlant, le nombre de leurs espèces étant supérieur à celui des mammifères. A ce jour, environ 700 espèces de dinosaures non-aviens (exclusion faite des oiseaux, donc) ont été découvertes. Un peu plus de la moitié d'entre elles ne sont connues que par un seul spécimen, et moins de 20% sont connues par plus de cinq spécimens. Cependant, un étude récente suggère que 71% des genres de dinosaures restent encore à découvrir, ce qui porterait à environ 1850 le nombre d'espèces de dinosaures ayant existé.
Premières découvertes et étymologie

En 1819 en Angleterre, Gideon Mantell (ci-contre à gauche), médecin et paléontologue, découvre par hasard les dents fossilisées d'un animal qu'il ne peut identifier.
La communauté scientifique de l'époque ne s'intéresse guère à cette
découverte, considérant qu'il s'agit de dents de poissons ou de mammifères. Mantell est cependant convaincu qu'il n'en est rien, et les rapproche plutôt des dents de l'iguane, en beaucoup plus grandes. Il estime ainsi la taille de l'animal à 20 mètres de long, et le nomme Iguanodon, c'est-à-dire "dent d'iguane".

Il faut cependant attendre 1842 pour que le terme dinosaure soit employé pour la première fois. C'est Richard Owen (ci-contre à droite), paléontologue anglais, qui propose lors de la "British Association of Sciences" le terme Dinosauria, dérivant de deux racines grecques, deinos, "effroyablement grand" ou "terrible", et sauros, "lézard" ou "reptile". Owen incluait alors dans ce groupe trois genres : Iguanodon, Megalosaurus et Hylaesaurus.
Voici donc nommés les "terribles lézards"...
Définition phylogénétique
Pendant très longtemps, les dinosaures ont été considérés comme des "reptiles géants", mais ces termes ne sont pas adéquats car, d'une part, le concept de "reptile" ne recouvre pas une réalité zoologique (ce n'est pas un groupe monophylétique acceptable car il regroupe plusieurs lignées n'ayant aucun ancêtre commun) et, d'autre part, tous les dinosaures n'étaient pas des géants. Certains ne dépassaient même pas la taille d'une poule ! De plus, les grands reptiles fossiles de l'?e secondaire n'étaient pas tous des dinosaures. Trop souvent encore les profanes, voire certaines revues de vulgarisation, ne distinguent pas les dinosaures de leurs contemporains plésiosaures, mosasaures, ichtyosaures et ptérosaures. Les dinosaures "vrais" sont exclusivement terrestres, voire arboricoles, mais certainement pas marins ou volants ! Pas d'ailes ni de palettes natatoires chez le dinosaure.
Tous les dinosaures se déplacent sur deux ou quatre pattes, lesquelles sont redressées bien à l'aplomb en dessous du corps, et non en position latérale comme chez les lézards et les crocodiles. Afin de distinguer les dinosaures de leurs plus proches parents (les dinosauromorphes), les scientifiques modernes, qui définissent aujourd'hui des groupes d'organismes d'après leur ancêtre commun et non sur la base du partage de certains caractères anatomiques, ont défini les dinosaures comme étant "les membres du clade le moins inclusif comprenant Triceratops horridus et Passer domesticus (le moineau domestique)". Cette définition phylogénétique ne peut être comprise qu'en utilisant un cladogramme, ou arbre phylogénétique, illustrant les liens de parenté entre les êtres vivants et les groupes au sein desquels ils se classent. Ainsi, est un dinosaure tout être vivant qui se positionne, dans un cladogramme, dans ce groupe d'organisme comprenant à la fois le Triceratops, le moineau et l'ensemble des descendants de leur ancêtre commun (le noeud de l'arbre où leurs deux branches se rejoignent). Toute espèce descendant de cet ancêtre commun (toujours considéré comme hypothétique en phylogénie) est un dinosaure.
Tous les dinosaures se déplacent sur deux ou quatre pattes, lesquelles sont redressées bien à l'aplomb en dessous du corps, et non en position latérale comme chez les lézards et les crocodiles. Afin de distinguer les dinosaures de leurs plus proches parents (les dinosauromorphes), les scientifiques modernes, qui définissent aujourd'hui des groupes d'organismes d'après leur ancêtre commun et non sur la base du partage de certains caractères anatomiques, ont défini les dinosaures comme étant "les membres du clade le moins inclusif comprenant Triceratops horridus et Passer domesticus (le moineau domestique)". Cette définition phylogénétique ne peut être comprise qu'en utilisant un cladogramme, ou arbre phylogénétique, illustrant les liens de parenté entre les êtres vivants et les groupes au sein desquels ils se classent. Ainsi, est un dinosaure tout être vivant qui se positionne, dans un cladogramme, dans ce groupe d'organisme comprenant à la fois le Triceratops, le moineau et l'ensemble des descendants de leur ancêtre commun (le noeud de l'arbre où leurs deux branches se rejoignent). Toute espèce descendant de cet ancêtre commun (toujours considéré comme hypothétique en phylogénie) est un dinosaure.
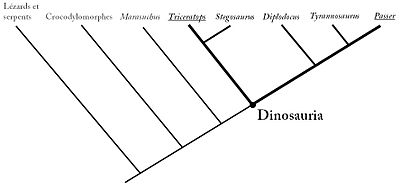
Schéma illustrant la définition actuelle du clade des dinosaures
Classification des dinosaures
Par la suite, les dinosaures ont été divisés en deux grands groupes : les saurischiens, "à bassin de lézard", et les ornithischiens, "à bassin d'oiseau".
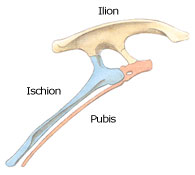
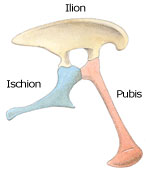
Bassin d'ornithischien (à gauche) et de saurischien (à droite)
Les saurischiens, aussi appelés sauripelviens, ont un bassin semblable à celui des autres reptiles comme les crocodiles et les lézards. Leur pubis, dirigé vers le bas et vers l'avant, forme un V à l'envers avec la partie inférieure arrière du bassin, l'ischion, l'illion formant la partie supérieure du bassin.
Cet ordre est divisé en deux sous-ordres :
→ les théropodes, c'est-à-dire les "pieds de bête sauvage", comprenant notamment tous les dinosaures bipèdes carnivores, mais aussi la familles de coelurosauriens comme l'Archeopteryx, famille probablement à l'origine des oiseaux modernes ;
→ les sauropodomorphes, ou "formes à pieds de lézard", comprenant essentiellement les quadrupèdes herbivores ayant un long cou et une grande queue, comme les diplodocidés, les brachiosauridés, les camarasauridés et les titanosauridés.

Saurischien et bassin de saurischien
Les ornithischiens ont un bassin plus proche de celui des oiseaux, sans pour autant avoir de lien de parenté avec eux. Chez ces dinosaures, les deux parties inférieures du bassin, le pubis et l'ischion, sont parallèles et dirigées vers le bas et l'arrière. Tous les ornithischiens étaient herbivores.
Cet ordre est divisé en trois sous-ordres :
→ les ornithopodes ou "pieds d'oiseau", possédant de nombreuses dents pour mastiquer les feuilles (jusqu'à 1000 dents jugales) ;
→ les thyréophores, "porteurs de bouclier", assez primitifs, aux mâchoires moins complexes que celles des ornithopodes et des marginocéphales. Leur dos était cuirassé par des plaques osseuses, des piques ou des éperons ;
→ les marginocéphales, "à tête ornée", de tailles très variées et regroupant des dinosaures comme les pachycéphalosauridés et les cératopsiens.
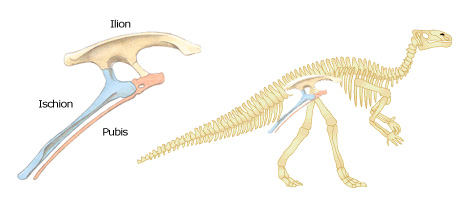
Ornithischien et bassin d'ornithischien
Les caractéristiques des dinosaures et les 9 homologies dinosauriennes
Chaque dinosaure possède ses propres caractéristiques. Cependant, certaines d'entre elles sont communes à l'ensemble du groupe :
→ chez les dinosaures, la tête articulaire du fémur est bien différenciée et déplacée vers l'intérieur du corps (position partagée aussi par les mammifères), permettant aux animaux de se déplacer en position verticale, les membres bien redressés sous le corps, contrairement aux reptiles primitifs et aux lézards et crocodiles, chez qui le fémur horizontal et les membres éloignés du tronc ne permettent pas de soulever nettement le corps ;
→ les dents des dinosaures étaient uniformes. Il n'y avait pas de distinction de type comme chez les mammifères, sauf exception ;
→ la ponte est le seul mode de reproduction connu à l'heure actuelle ;
→ une fenêtre osseuse crânienne supplémentaire, située devant l'orbite et bien distincte et séparée de celui-ci, appelée fenêtre antéorbitaire et distinguant les dinosaures des autres reptiles ;
→ un coeur à quatre cavités, pompant le sang en deux étapes à partir d'une aorte unique, comme chez les mammifères et les oiseaux mais très éloigné de ce qu'on retrouve chez les reptiles modernes. Cela suggère l'existence d'un système respiratoire et d'un système circulatoire séparés, permettant un métabolisme plus rapide que celui des animaux à sang froid. Il est donc possible que les dinosaures aient eu le sang chaud.
Cependant, ces caractéristiques laissent encore trop d'exceptions et d'incertitudes. Mais il est possible de pallier ce problème en s'appuyant sur d'autres caractéristiques des dinosaures. Ainsi, qu'il soit ornithischien ou saurischien, chaque dinosaure présente, en principe, une série de 9 homologies (héritées d'un ancêtre commun, ce sont des liens évolutifs entre deux traits observés chez deux espèces distinctes).
Homologie n°1 : les vomers (os du palais) sont très allongés et atteignent voire dépassent le bord antérieur de la fenêtre antéorbitaire. Il s'étendent u bout du museau jusqu'à cette cavité.
→ chez les dinosaures, la tête articulaire du fémur est bien différenciée et déplacée vers l'intérieur du corps (position partagée aussi par les mammifères), permettant aux animaux de se déplacer en position verticale, les membres bien redressés sous le corps, contrairement aux reptiles primitifs et aux lézards et crocodiles, chez qui le fémur horizontal et les membres éloignés du tronc ne permettent pas de soulever nettement le corps ;
→ les dents des dinosaures étaient uniformes. Il n'y avait pas de distinction de type comme chez les mammifères, sauf exception ;
→ la ponte est le seul mode de reproduction connu à l'heure actuelle ;
→ une fenêtre osseuse crânienne supplémentaire, située devant l'orbite et bien distincte et séparée de celui-ci, appelée fenêtre antéorbitaire et distinguant les dinosaures des autres reptiles ;
→ un coeur à quatre cavités, pompant le sang en deux étapes à partir d'une aorte unique, comme chez les mammifères et les oiseaux mais très éloigné de ce qu'on retrouve chez les reptiles modernes. Cela suggère l'existence d'un système respiratoire et d'un système circulatoire séparés, permettant un métabolisme plus rapide que celui des animaux à sang froid. Il est donc possible que les dinosaures aient eu le sang chaud.
Cependant, ces caractéristiques laissent encore trop d'exceptions et d'incertitudes. Mais il est possible de pallier ce problème en s'appuyant sur d'autres caractéristiques des dinosaures. Ainsi, qu'il soit ornithischien ou saurischien, chaque dinosaure présente, en principe, une série de 9 homologies (héritées d'un ancêtre commun, ce sont des liens évolutifs entre deux traits observés chez deux espèces distinctes).
Homologie n°1 : les vomers (os du palais) sont très allongés et atteignent voire dépassent le bord antérieur de la fenêtre antéorbitaire. Il s'étendent u bout du museau jusqu'à cette cavité.

 Observation des vomers sur un crâne de Tyrannosaure rex
Observation des vomers sur un crâne de Tyrannosaure rexHomologie n°2 : tous les dinosaures présentent un sacrum, c'est-à-dire un ensemble de vertèbres soudées situé au-dessus de l'illion. Pour qu'une espèce soit considérée comme un dinosaure, il faut que son sacrum soit constitué d'au moins trois vertèbres soudées. Il peut donc y en avoir plus selon les espèces.
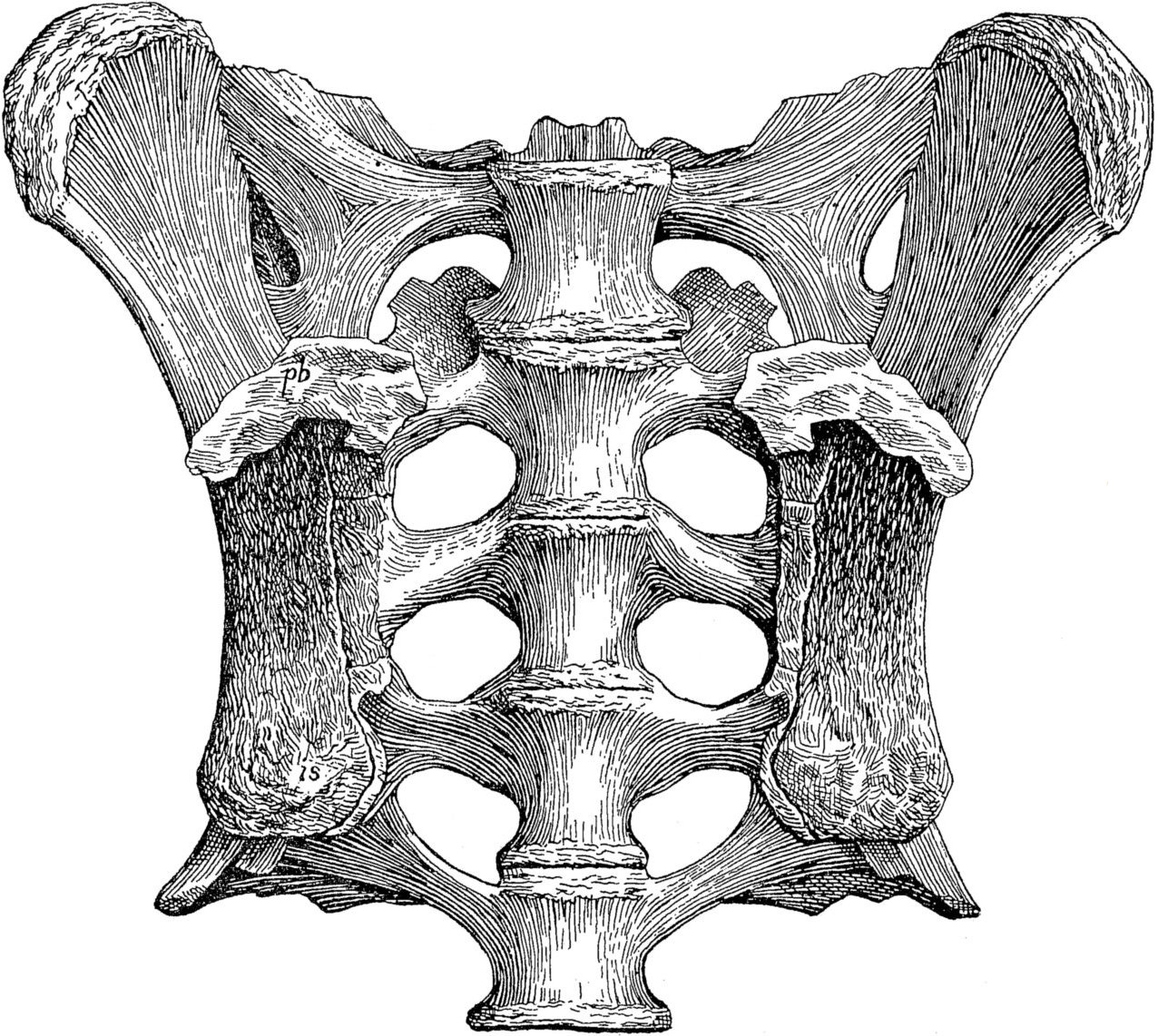
Sacrum d'Iguanodon (en bleu) : 7 vertèbres soudées

Sacrum de Stégosaure : Sacrum de Brachiosaure :
4 vertèbres soudées 5 vertèbres soudées
Homologie n°3 : la cavité glénoïde, cavité articulaire de l'épaule recevant la tête de l'humérus, formée par la scapula (omoplate) et l'os coracoïde, est tournée vers l'arrière chez les dinosaures, contrairement aux crocodiliens chez qui elle est positionnée latéralement.

Homologie n°4 : l'humérus doit posséder une crête delto-pectorale, permettant l'insertion des muscles releveurs du bras, laquelle doit mesurer entre un tiers et la moitié de la longueur du corps de l'humérus.

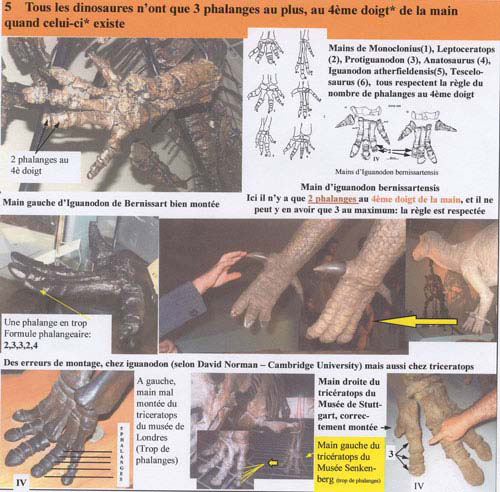
Homologie n°5 : le quatrième doigt de la main, s'il existe, ne doit posséder que trois phalanges. Il arrive, dans certains musées, une phalange en trop, rajoutée après coup par les scientifiques qui pensaient que l'os n'avait tout simplement pas été retrouvé.
Homologie n°6 : l'acétabulum, ou cavité acétabulaire, qui constitue l'articulation de la hanche et reçoit la tête du fémur latéralement, est largement ou complètement ouvert. De plus, la crête supra-acétabulaire est proéminente et empêche tout mouvement latérale de la patte, agissant comme un cran d'arrêt.


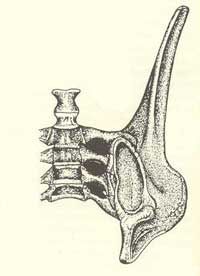
Homologie n°7 : la tête du fémur doit être en forme de boule et complètement déjetée, avec un col du fémur bien distinct.

Articulation de la hanche chez le Diplodocus
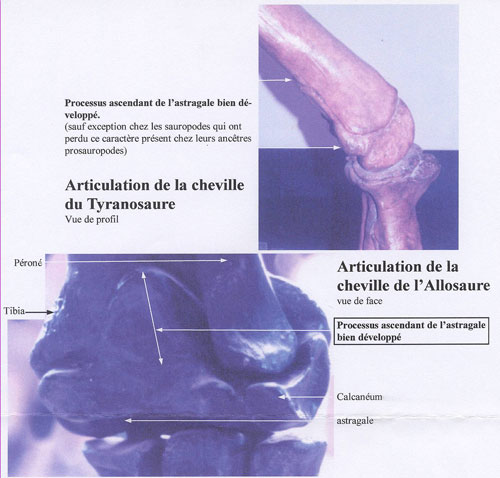
Homologie n°8 : le processus ascendant de l'astragale est développé. Attention cependant, ce caractère est absent chez les sauropodes, qui auraient perdu cette homologie, cependant présente chez leurs ancêtres prosauropodes. Autre remarque : le processus de l'astragale est très développé chez l'Archeopteryx, ce qui le rapproche des dinosaures. Est-il alors un oiseau ou un dinosaure ?
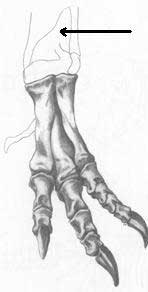


Chez le Tyrannosaure Chez le Triceratops Chez l'Archeopteryx
Homologie n°9 : le péroné est réduit par rapport au tibia, même si cela est moins marqué chez les sauropodes du fait de leur poids.
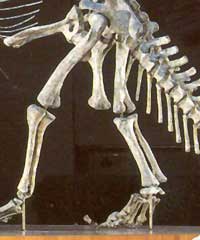

Chez l'Iguanodon (à gauche) et le Stégosaure (à droite)
Sources : Wikipedia, Encyclopédie Universalis, jurassic-world.com, livre "Grandeur et décadence des dinosaures" (Philippe Tacquet)
Afficher la suite de cette page